The editor’s choice for our July issue is “Consistency of demographic trade-offs across 13 (sub)tropical forests” by Kambach et al. Here, Associate Editor Pieter Zuidema explains the importance of this research:
There is no such thing as ‘the tropical forest’. Tropical forests differ in their structure, diversity and species composition. These differences are caused by variation in climate, biogeographical region and disturbance regimes. For instance: Asian forests are generally taller than African forests, Caribbean forests are more prone to hurricane damage while others are not, and tree species composition is very distinct across tropical continents. In spite of these striking differences across forests, tropical tree species seem to have adopted similar sets of life history strategies to live and survive in these ecosystems. Those strategies align along two major axes of variation. One axis that separates species that grow fast but have little tolerance to survive under shade, from those that tolerate shade but grow slowly. And another axis that distinguishes species reaching large sizes, from those that produce abundant offspring. This, in short, is the conclusion of the paper by Stephan Kambach and co-workers “Consistency of demographic trade-offs across 13 (sub)tropical forests”.
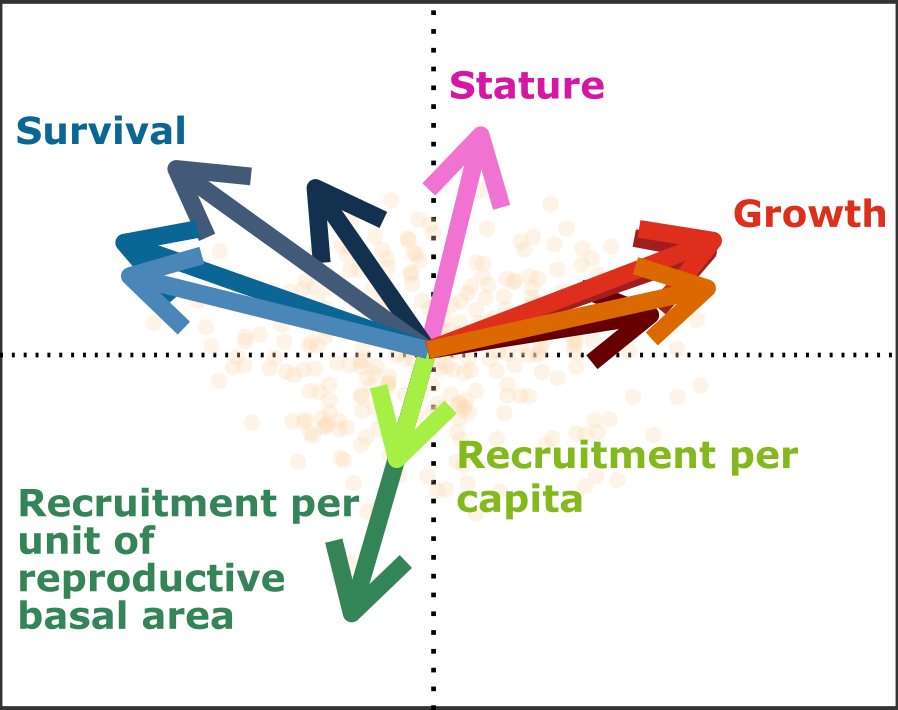
The growth-survival axis is a longstanding phenomenon for tropical tree species and plants in general, but the second axis – recruitment vs. maximum size – is much more poorly studied. The important contribution of the Kambach et al. paper is that evidence for these axes was assessed for 13 (sub-)tropical forest sites varying in climate, structure, species composition and disturbance regime. These sites ranged from deciduous to evergreen, species-rich to hyper-rich and from highly to mildly disturbed. Across sites, and using demographic data from an impressive amount of tree species (5 thousand) and individuals (2.5 million), the authors were able to consistently detect these two axes of life history strategies for 12 out of 13 sites. To get to these results, the authors calculated species-specific rates of growth, survival and recruitment. An important novelty is that these demographic rates were calculated per canopy layer, which allowed comparing results across species differing in maximum height, and forests differing in canopy height. They then combined demographic rates for all species in a principle component analysis and checked for significant associations of rates with the two first axes. Opposite and significant alignment of survival vs. growth and of recruitment vs. maximum size along the two main components were considered as support for the two life history axes.
As a final step, the authors combined site-specific results to check for differences between sites. This showed that demographic strategies are quite similar across plots, but also revealed some differences between Asian and African sites, and between the most hurricane-prone site compared to the rest. Thus, while those two main axes of life history variation dominate, differences across forests modify those axes. Possible next steps in the analyses of tropical tree life histories are studies on the phylogenetic signature and on the implications of strategies for population dynamics.
For vegetation modellers, this study contains some good news: in spite of the huge diversity of species and the variety of forest types, tropical forest dynamics may be fairly well represented by a limited number of plant functional types.

