Eric Griffin and colleagues recently had their paper, Plant host identity and soil macronutrients explain little variation in sapling endophyte community composition: Is disturbance an alternative explanation?, published in Journal of Ecology. You can read more about the paper below.
Recent advances in high-throughput sequencing are enabling scientists to uncover the structure and function of microbial communities, which many have called “The Great Unseen” component of ecosystems. As previous studies of the human gut microbiome have demonstrated, plants – like humans – host an abundant and diverse array of microbes on (“epiphytes”) as well as inside them (“endophytes”) which function as critical mediators of host performance (Vandenkoornhuyse et al. 2015, Griffin et al. 2016, 2017). Indeed, it is becoming increasingly clear that microbial communities are important components of plant function as well as entire plant community dynamics (e.g., Mangan et al. 2010, reviewed by Griffin & Carson 2015).

Figure 1: (left) Sorocea affinis sapling, one of the five tree species used in the study, (right): Alseis blackiana sapling, one of the five tree species used in the study.
In this study, we used next-generation sequencing (Illumina MiSeq) to characterize bacterial endophyte community composition among saplings of co-occurring tree species (Figure 1) in a tropical forest in Panama, where microbial growth and persistence are likely high. We tested a series of hypotheses to understand whether host species, nutrient availability, and disturbance (via commercial antibiotic applications) are most important in determining variation in endophyte community composition. Moreover, we tested whether a “core microbiome” occurred among all tree species, whereby a small subset of bacterial taxa occurred on almost all saplings from all study species. To test these hypotheses, we used a long-term forest fertilization experiment on the Barro Colorado Nature Monument (Gigante Penninsula, Figure 2) where nitrogen, phosphorus, and potassium and all of their combinations had been added to soil for over 10 years. In addition, we artificially disturbed one of our focal species by applying antibiotics for almost three years within the soil (N, P, K) fertilization experiment.

Figure 2: Barro Colorado Island, part of the Barro Colorado Nature Monument, Panama.
To our surprise, host identity and soil nutrient additions explained little (<10%) variation in endophyte community composition (Figure 3a-c), a finding which is in stark contrast to studies of bacterial epiphytes, fungal epiphytes, and insect herbivores, which all show relatively high levels of host specificity (e.g., Kembel et al. 2014; Kembel & Mueller 2014; Forister et al. 2015). Conversely, we found clear evidence for a “core microbiome,” where nine bacterial taxa were present on almost all individuals of all five tree species and made up almost a third of all total bacterial sequences. Lastly, disturbance (antibiotics) explained more variation in endophyte community composition compared to all nutrient addition combinations combined and twice the variation explained by host identity for all five tree species (Figure 3d-f).
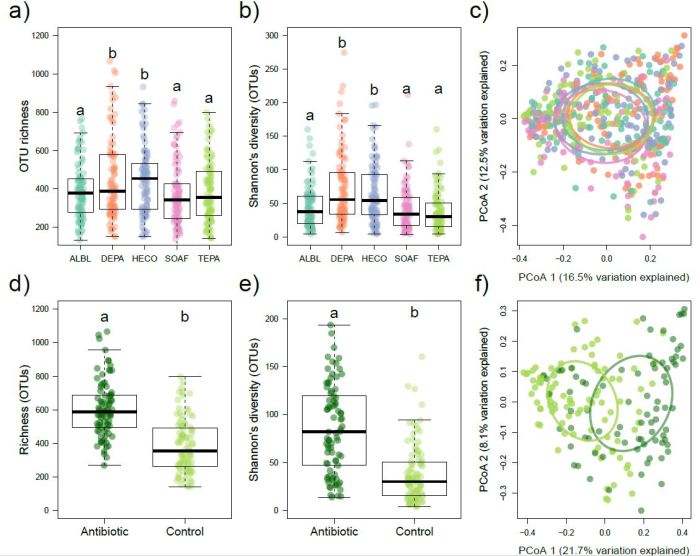
Figure 3: Differences in bacterial endophyte richness (a) and diversity (b) among saplings of Alseis blackiana (ALBL), Desmopsis panamensis (DEPA), Heisteria concinna (HECO), Sorocea affinis (SOAF), and Tetragastris panamensis (TEPA). Letters denote significant pairwise differences (P < 0.05) between tree species as determined by mixed effects analysis. (c) Principle component analysis (PCoA) of endophyte communities. Colors correspond with those in panels a and b. Ellipses denote 95% standard deviation from the centroid for each species. Effects of foliar antibiotic applications on richness (d) and diversity (e) of TEPA. Letters denote significant differences as determined via mixed effects modelling. (f) PCoA of bacterial endophyte community of TEPA. Colors denote antibiotic treatment status (light green = control; dark green = antibiotics) and correspond to those in panels d and e.
Overall, our results challenge the idea that host identity is a primary filter shaping bacterial endophyte communities and suggest that many of the same bacterial taxa occur inside plant hosts even if those host plants are phylogenetically diverse. Moreover, we found that disturbance was an important driver of microbial community composition, which parallels the importance of disturbance in other areas of community ecology.
Eric Griffin (Smithsonian Environmental Research Center, USA) and colleagues.
Read the full paper online: Plant host identity and soil macronutrients explain little variation in sapling endophyte community composition: Is disturbance an alternative explanation?



